231004投稿
Hsp90とepigenetic因子の共同作業:Sollarsらの研究結果(2003年)
Hsp90が、変異遺伝子による異常形態の発現を抑圧するメカニズムとして、第一に、タンパク質レベルのbuffering反応が考えられる。すなわち、Hsp90が分子シャペロン効果を発揮して、その変異標的タンパク質の構造上および機能上の異常を緩和(広義の修復)する場合である。第二は、変異遺伝子の発現を抑制する場合である。ただし、後者の場合、変異遺伝子と正常遺伝子を区別してそれらの発現(例えば、転写)を抑制することはできない。上述したR&Lの研究では第一のケースを想定していた(HafenらのRaf kinaseの研究が念頭にあった(文献13))。ところが、第二のケースを強く示唆する結果が、Sollarsらによって示された(文献20)。
- Sollars, V. et al. Nature Genet. 33: 70 (2003)
第一の遺伝子変異に基づく形態異常を増幅する第二の遺伝子変異の同定
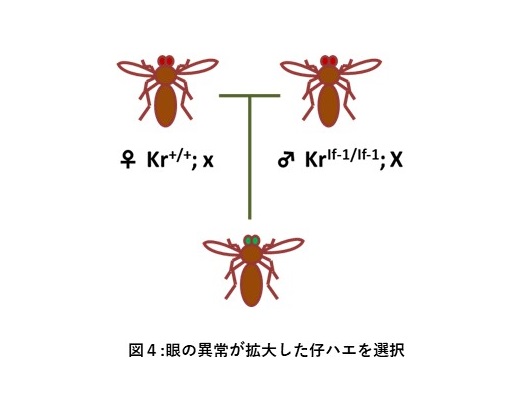
ショウジョウバエのKruppel(Kr)遺伝子は胸・腹部の体節形成を支配する転写因子Krをコードしており、KrIf-1変異はKrタンパク質の異所性発現(outgrowth)を引き起こす優性変異である。しかし、KrIf-1変異単独では、眼のサイズが小さくなるなどの軽微な変異表現型を示すにすぎない。Sollarsらは、deficiency chromosome screening法によって、KrIf-1の異常表現形質を増幅し、眼の顕著な異所性発現をもたらす第二の変異遺伝子群をスクリーニングした(図4)(文献20)。その結果、9種類のtrithorax group(TrxG)関連遺伝子の変異およびHsp83遺伝子の変異5種類を同定した。このKrIf-1変異増幅結果は、第二の変異遺伝子がヘテロの♀とKrIf-1のホモの♂を掛け合わせたときに認められ、♂と♀を逆にしたクロスでは形態異常を示す子は得られなかった。つまり、これらの変異遺伝子の母系効果によって、眼の異所性形成増幅が起こったことになる。