230908 投稿
R&Lによる選択形質の継代実験
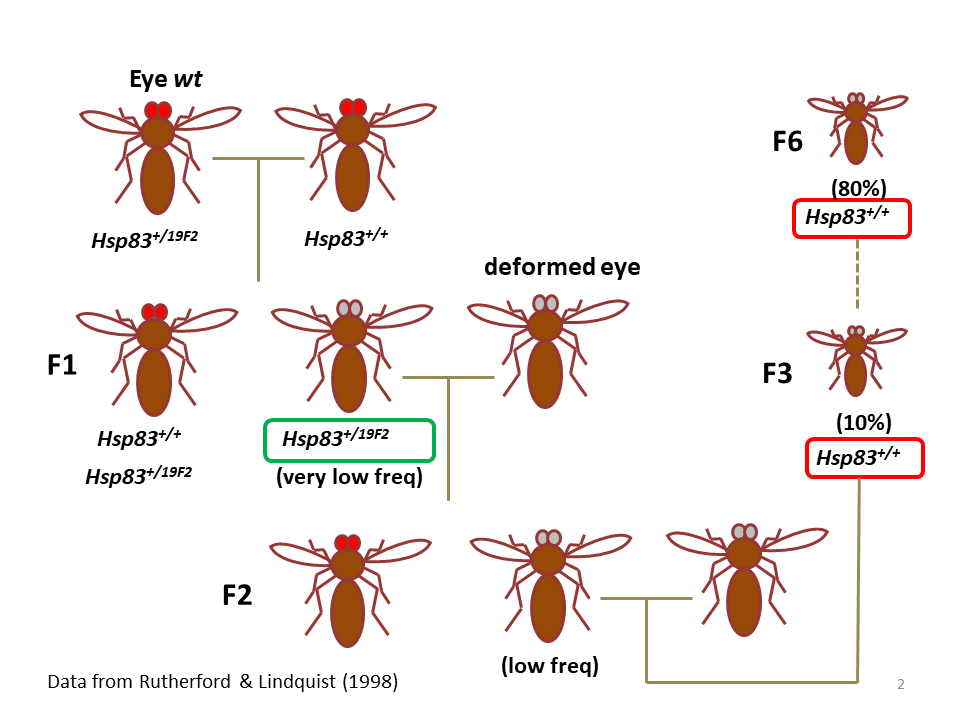
交配実験例を見てみる(図2)。変異Hsp83(19F2)をヘテロにもつハエとラボ株Hsp83(+)とをクロスし、眼が形態異常を示すF1♂を選んだ。このF1♂を同じクロスから生まれた形態が正常なF1♀(Hsp83 19F2/+)とクロスして、F2を作った。F2にはF1♂と同じように眼が異常なものが低頻度の割合で含まれていた。選択実験は、形態異常のF1同士をクロスしてF2を作ること(HE系)、また形態正常のF1どうしをクロスしてF2を作ること(LE系)によって行った。14継代クロスして樹立したHE系では、90%に至る割合のハエが眼の形態異常を示したが、LE系では形態異常のハエはほとんど生まれなかった。HEとしてHE1, HE2, HE3の3系統を作ったが、眼の形態異常の特徴と程度が3系統間で違っていた。つまり、ラボ株で隠されていた眼の形態異常をもたらす「変異遺伝子」は複数種類あることが示された。なお、次項でくわしく論ずるが、前のセンテンスの「変異遺伝子」は、正確には「変異遺伝子セット」とすべきである。
ここで、R&Lは不可解な事実に直面した。上記のHE株は、現れたハエの眼の異常形態を指標にして選択を繰り返し行って作成した。彼女たちの解釈によれば、異常表現型のハエは、Hsp83+/19F2が形態異常の基となる変異遺伝子の表現型への変換をbufferingしきれないので、出現したのである。ところが、樹立したHE株において、異常表現型を示すハエはHsp83(19F2)を持っていなかった(Hsp83+/+)。HE株に、さらに2コピーの野生型Hsp83遺伝子を導入しても、形質は依然として異常表現型であった(十分量のHsp83が存在しても、変異は抑制できない)。また、HE株のハエについて、Hsp83遺伝子の塩基配列を調べたところ、どれも野生型Hsp83であり、Hsp83(19F2)の配列(Arg48⇒Cys)はなかった。すなわち、継代によって変異Hsp83依存的に発現した形態異常が変異Hsp83依存性を示さなくなった。言い換えれば、Hsp83がホモ野生型であっても、形態異常の基となる遺伝子変異がbufferingされなくなった。すなわち、Waddingtonが上記の2つの研究において観察した、継代によるフェノコピーの刺激応答性がなくなり、刺激非応答的な形態異常の発現への変換によく似た結果が、この実験でも認められたのである。