230910 投稿
多重遺伝子変異が原因か?
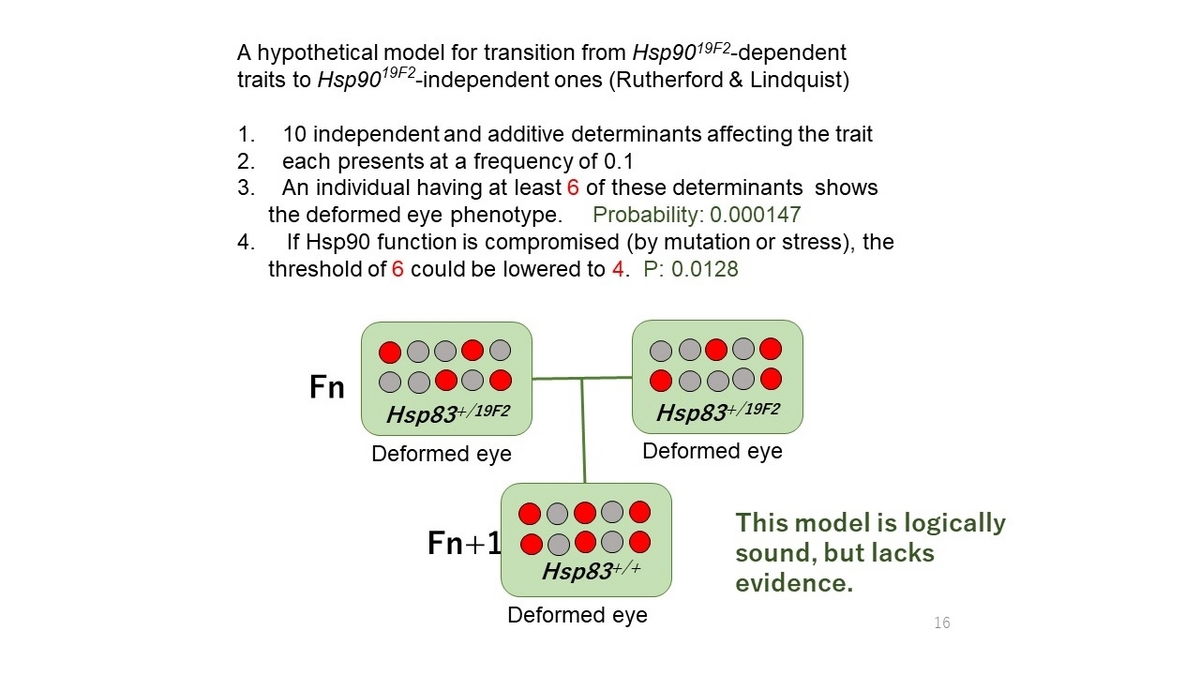
HE株において、「異常表現型の基となる変異遺伝子がHsp83によるbufferingを受けないように変化した」、とはどういうことなのか。この問題にたいして、Lindquistたちは、多重責任遺伝子仮説による説明を考えた。以下、いささかくどい説明になるが、彼女たちの論文の記述にしたがう。まず、問題としている形質(眼の形態)にかかわる遺伝子が10個あると仮定する。それぞれの遺伝子には眼の異常の原因となるalleleが0.1の割合で存在していると仮定する。そこで、問題としている眼の異常形態が発現するためには、10個のうち最低6個の遺伝子の変異allele型を必要と仮定する(それぞれの変異alleleは等価であるとする)。言い換えれば、異常形態発現の閾値が変異allele型 6種と仮定した。すると単純な組み合わせ問題の計算によって、この集団では、7,000匹に1匹のハエの目が異常形態になる。もし、Hsp83変異によってHsp83のbuffering機能が低下すると、異常形態発現の<閾値>が下がって、最低4種の遺伝子の変異allele型で眼の異常表現型が現れると仮定する。この条件では、78匹中1匹のハエが眼の異常を示すはずである。HE株の作製には、最初はHsp83 19F2/+の異常形質を示しているハエ(最低4種の変異allele型を有する。変異allele型は図では赤丸でしめしてある)を選択して継代交配した。上述したように、F1以後の継代交配においては、眼の異常形質を指標として親を選び交配させた。つまり、どちらの親ハエも、少なくとも4種の変異allele型を持っていることになる。このやり方だと、継代を続けると、交配に使う親ハエのもつ変異allele型は増えると予想される(ただし、近親交配なので、最初に使った♂と♀のハエが持つ変異allele型の総数を超えることはない)。例えば、仮に5代目の継代交配に使ったハエが6個の変異allele型をもつとすれば、そのハエはHsp83(+/+)でも形態異常でありうる(図3)。かくして、継代を続けるにしたがって、形態異常を示すハエのもつallele型の数が増し、Hsp83(+/+)の割合が増えてくるので、HE株がHsp83(+/+)であっても不思議ではない。とりわけ、Hsp83(+/+)の個体がHsp8319F2/+の個体よりも、少しでも繁殖優位性を持つならば(十分にありうる)、樹立したHE株がHsp83(+/+)である可能性は高いはずである。
上記の計算で仮定した、Hsp83のbufferingを受ける仮定の10種の変異allelesを実験的に決めることは、現代の先端実験技術といえども煩雑すぎて手におえない(全ゲノム塩基配列決定がさらに簡便になれば不可能ではないが、やる価値があるかどうか不明)。さらに、変異表現型が出現するための<閾値>は単なる概念でしかなく、強いて言えば、epigeneticな機構の一つとしか言えない。それ故、多重遺伝子変異仮説は、多重遺伝子変異の同定と、<閾値>を決めるメカニズムの解明を必要としている。
なお、R&Lの研究では、Waddingtonの研究のように、変異表現型の責任遺伝子変異が優性の単独遺伝子変異であるというような結果は得られていない。